EL FLUJO DE ENERGÍA EN LOS ECOSISTEMAS
Martín Oesterheld, María Semmartin y Mariana Ciavattini

Índice
La visión del ecosistema
Cuando en los capítulos sobre interacciones entre poblaciones te hacías preguntas como “¿Qué efecto tiene un aumento de la densidad de herbívoros?”, las respuestas apuntaban a lo que podía pasar con la abundancia de ciertas especies de plantas, de ciertos depredadores, o de ciertos competidores de esos herbívoros. Pero lo visto en esos capítulos no te servía de mucho para responder qué efecto tiene un aumento de la densidad de herbívoros sobre la frecuencia de incendios, o sobre la llegada de carbono al suelo, o sobre la productividad.
Estas nuevas preguntas requieren una visión diferente, un marco de referencia que va más allá de las interacciones entre poblaciones: una visión de ecosistema. Esta visión considera a las interacciones entre poblaciones como parte de un sistema más complejo, compuesto por cientos o miles de poblaciones que hacen fluir la energía y circular los nutrientes. Por ejemplo, una visión ecosistémica podría considerar que si la densidad de herbívoros aumenta, la tasa de descomposición de la materia orgánica podría modificarse porque cambia la cantidad y la calidad de la hojarasca que llega a los microorganismos del suelo. Otro ejemplo: si la densidad de herbívoros disminuyera, la frecuencia de incendios podría aumentar porque se acumularía más pasto muerto inflamable. Estos cambios, a su vez, podrían alterar los ciclos de nutrientes y afectar a la estructura de las comunidades vegetales. No es que los modelos de Lotka-Volterra vistos en la primera parte del libro sean irrelevantes. Se trata de otra visión, como ejemplifica esta obra de arte.

Imagen 1. Ejemplo de visión ecosistémica: Cambios de densidad de herbívoros pueden alterar la frecuencia de incendios de un pastizal
Si vas a aprender sobre una visión de ecosistema, es oportuno definir a los ecosistemas y hablar de sus límites. Un ecosistema es un conjunto de organismos vivos y su entorno físico, interconectados mediante un flujo de energía y un ciclo de materiales (Tansley 1935, aquí puedes ver más detalle sobre este concepto). ¿Dónde empieza y termina un ecosistema? La vida está interconectada a nivel global y, por lo tanto, en definitiva hay un solo ecosistema: el planeta. Sin embargo, frecuentemente reconocemos subsistemas dentro de ese ecosistema global y hablamos, por ejemplo, del ecosistema de tal río o tal bosque, a los cuales les asignamos límites. En algunos casos, estos límites te pueden resultar naturales y evidentes, como un lago o una isla. En otros casos, tendrás que definirlos más o menos arbitrariamente según las preguntas que quieras responder. Si te interesa, por ejemplo, el impacto de una actividad agrícola, podrías considerar que el ecosistema corresponde a un lote o potrero y “termina en el alambrado”. Esa decisión no es trivial. Puede ser muy acertada para ciertas preguntas (por ejemplo, ¿aumenta el riego la radiación solar absorbida por el lote?) y engañosa para otras (por ejemplo, ¿qué impacto tiene una fertilización excesiva de este lote?). En este último caso, la fertilización puede ser bastante inocua si el ecosistema se limita al lote, pero perjudicial si se incluyen cuerpos de agua vecinos.

Imagen 2. Las lagunas de Epulauquen en el norte de Neuquén pueden ser consideradas como dos ecosistemas, como uno o como parte de un ecosistema mayor.
Este capítulo aborda el flujo de energía dentro de los ecosistemas. En primer lugar, estudiaremos cómo la energía ingresa a los ecosistemas, pasa de un nivel tróficoPosición que ocupa un organismo en la cadena alimentaria de un ecosistema. a otro y es disipada por todos ellos. Segundo, profundizaremos en la complejidad de cada uno de esos niveles tróficos. Tercero, analizaremos el efecto de variables externas, muchas veces accionadas por humanos, que agregan energía con diversos fines. Cuarto, aprenderemos sobre la eficiencia energética de diferentes procesos ecosistémicos. Finalmente, estudiaremos la diversidad de variantes que tiene el flujo de energía entre tipos de ecosistemas.
Tipos de variables: estado, flujo y control
En el flujo de energía conviene distinguir entre variables de estado, de flujo y de control (Figura 1). Las variables de estado representan el tamaño de un componente y suelen dibujarse como rectángulos o “cajas”. En la figura, la variable de estado “consumidores primarios” corresponde a la cantidad de energía en ese nivel trófico. Sus unidades pueden ser, por ejemplo, MJ/haMegajoule o Megajulio. Las variables de flujo representan la velocidad de un proceso, se dibujan como flechas o cañerías y sus unidades incluyen al tiempo (por ejemplo, MJ/ha.día). En la figura, las dos variables de flujo representan la velocidad o ritmo con que los consumidores primarios consumen energía (izquierda) y la disipan por la vía de la respiración (derecha). Si estos fueran los únicos flujos relevantes, de un tiempo a otro, la magnitud de la variable de estado puede cambiar de acuerdo con el balance entre las entradas y las salidas. Finalmente, las variables de control representan efectos sobre un flujo o una caja y suelen dibujarse como flechas punteadas que actúan sobre una válvula. Por ejemplo, la temperatura puede afectar al consumo y a la respiración. Por lo tanto, el tamaños de la variable de estado cambiará de acuerdo con la temperatura y la forma particular en la que afecta a cada flujo.
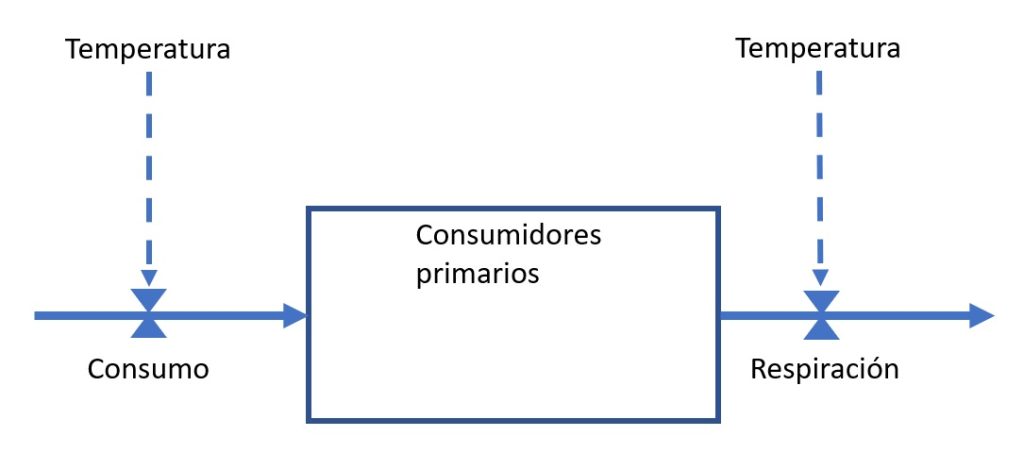
Figura 1. Variables de estado, de flujo y de control.
Un modelo genérico del flujo de energía
La fuente inicial de energía para todos los ecosistemas es el sol La cantidad de energía es enorme: unos pocos segundos de la energía solar que llega a la Tierra equivalen al consumo eléctrico mundial de un año. . (Hay otras fuentes iniciales de energía, pero son mínimas comparadas con el sol). En un ecosistema, los productores primarios son los únicos organismos capaces de incorporar directamente la energía solar a su metabolismo. En cambio, los consumidores sólo pueden incorporar a su metabolismo energía contenida en productores primarios o en otros consumidores que se suceden a lo largo de la cadena trófica. Finalmente, los descomponedores consumen energía contenida en material muerto. Considera un ecosistema genérico, con cinco variables de estado que corresponden a la energía contenida en los productores primarios, en los consumidores primarios y secundarios, en los descomponedores y en la necromasa, que es la materia orgánica muerta (Figura 2). Hacia y desde cada uno de ellos, la energía fluye según variables de flujo que describen procesos. Como puedes ver, estas variables de flujo no están representadas por flechas sino por cañerías que cumplen la misma función, pero además transmiten una idea de su magnitud a partir del ancho. Las variables de control aparecerán más adelante para mantener simple la figura por ahora.

Figura 2. Flujo de energía en el ecosistema. Las cajas representan reservorios de energía en niveles tróficos (línea punteada), en la necromasa (línea gris), y en el ecosistema (línea negra llena). El sistema de cañerías representa flujos de energía entre reservorios. Niveles tróficos: P = productores, C1 = consumidores primarios, C2 = consumidores secundarios, D = descomponedores. Flujos: RI = radiación fotosintéticamente activa incidente, NU = no utilizado, RA = radiación absorbida, NA = no asimilado, R = respiración, PPB = productividad primaria bruta, PPN = productividad primaria neta, C = consumo, PSB = productividad secundaria bruta, PSN = productividad secundaria neta, M = muerte, PBD = productividad bruta de los descomponedores, PND = productividad neta de los descomponedores. Las cañerías verdes representan flujos de energía radiante, las rojas energía calórica y las azules energía química.
De la energía solar incidente total, aproximadamente la mitad llega en longitudes de onda que pueden ser captadas por los pigmentos fotosintéticos, la llamada “radiación fotosintéticamente activa” (RI en la Figura 2). La RI puede seguir esencialmente dos caminos: ser absorbida por los pigmentos fotosintéticos de los productores primarios (radiación fotosintéticamente activa absorbida, RA), lo cual en sistemas terrestres está directamente determinado por la cantidad de área foliar y la concentración de clorofila por unidad de área foliar, o ser absorbida por el suelo, o reflejada por la vegetación o el suelo, flujos que se engloban bajo el nombre de no utilizado (NU).

Imagen 3. En esta imagen de lotes agropecuarios del sur de la provincia de Buenos Aires puedes ver fuertes variaciones de la proporción de radiación incidente absorbida por los productores primarios. ¿Puedes indicar dónde es alta y baja e inferir las variables que la controlan? Fuente: Google Earth.
A su vez, parte de la radiación absorbida se transforma en fotosintatos y conforma la productividad primaria bruta (PPB). La PPB resulta de que los fotones absorbidos (RA) se transforman en ATP y NADPH en la fase lumínica de la fotosíntesis. Esas moléculas entregan energía para unir el CO2 a otras moléculas carbonadas en la fase química de la fotosíntesis. Ambos procesos son ineficientes y por lo tanto parte de la radiación absorbida (RA) no es asimilada (NA) y se pierde como calor, mientras que el resto constituye la productividad primaria bruta. Parte de ella es respirada (R) en el metabolismo de los productores primarios y el resto se incorpora a su biomasa a una tasa que se llama productividad primaria neta (PPN). Este es uno de los flujos más importantes en todo ecosistema ya que representa la energía capturada por los productores primarios que estará disponible para el resto de los niveles tróficos (Figura 2). Puedes verlo en acción en una una sola planta, a nivel ecosistema o a nivel global.
Aquí puedes ver más detalles sobre la respiración.

Imagen 4. En el día de la foto, las plantas de esta estepa patagónica absorbieron una parte de la radiación incidente (¿puedes estimar la proporción?). Parte de esa radiación fue transformada en fotosintatos (PPB). Como parte de su metabolismo, usaron una parte de su energía en la respiración (R). Otra parte de la energía quedó almacenada en el crecimiento de hojas, tallos, polen, raíces, etc. Esta última parte fue la PPN del día. Foto: Oesterheld.
Una parte de la PPN es consumida por los consumidores primarios (C) y el resto no es consumido y pasa a formar la necromasa (“no utilizado“, NU): los tejidos vegetales no consumidos senescen y se incorporan al suelo. Una parte de lo que consumen los consumidores primarios es asimilado y constituye la producción secundaria bruta. Es equivalente a lo que se transfiere a la sangre en los vertebrados y a la hemolinfa en los insectos . El resto es la energía no asimilada y se pierde en forma de excreciones (NA). Las bacterias y hongos que atacan plantas vivas también son consumidores primarios. Si bien no poseen ni sangre ni hemolinfa, se puede considerar que no asimilan todo lo que consumen. De la energía asimilada, los consumidores primarios destinan una parte a los procesos de mantenimiento y crecimiento. Ese consumo de energía está representado en la Figura 2 como pérdidas por respiración (R) y la energía remanente en los tejidos es la productividad secundaria neta (PSN), la cual queda disponible para los consumidores secundarios. El pasaje de energía es cualitativamente similar al descripto desde los productores a los consumidores primarios, ya que existe una porción no utilizada, otra no asimilada y otra respirada. Eventualmente, podría haber consumidores terciarios, etc., que no han sido dibujados por simplicidad y sólo se ha considerado que los secundarios mueren por causas distintas de la depredación (M). La necromasa puede ser aprovechada por los descomponedores con un esquema similar al descripto para los otros consumidores, con la salvedad de que al no existir una “productividad de la necromasa”, no hay un flujo de energía no utilizado por los descomponedores, los cuales, según el modelo, consumen directamente de la caja.
Una vez que has visto al detalle cada bifurcación del flujo de energía, te recomendamos que retomes la visión ecosistémica y prestes atención al sistema como un todo. Además de los organismos de los distintos niveles tróficos, están los factores abióticos y las múltiples relaciones entre todos. Una vez que toma la forma de productividad primaria bruta, la energía pasa a estar asociada con moléculas orgánicas compuestas por nutrientes que, como verás más adelante, juegan un papel crítico como variables de control del flujo. Por ejemplo, la caja de “necromasa” en la Figura 2 es estrictamente una cantidad de energía, pero esa energía está en forma de tejidos o células muertas o incluso moléculas orgánicas disueltas. Los nutrientes que contienen (nitrógeno o fósforo, por ejemplo) aparecerán en párrafos siguientes como controladores del flujo de energía.
Aquí puedes ver más detalles sobre la diferencia entre flujo y balance de energía.
No es lo que parece: preconceptos sobre los niveles tróficos
Seguramente al leer la sección anterior has asociado a los productores con plantas, a los consumidores primarios con paradigmáticos herbívoros y a los secundarios con clásicos depredadores. Sin embargo, los niveles tróficos no siempre se alinean de forma precisa con las divisiones taxonómicas de los organismos. Tomemos, por ejemplo, a la babosa de mar Elysia chlorotica, que puede “secuestrar” cloroplastos de las algas que consume y realizar fotosíntesis. En este caso, un consumidor primario actúa también como productor primario. En otro orden, hay plantas como Cuscuta sp., que carecen de clorofila y obtienen toda su energía al parasitar a otras plantas. Aunque son taxonómicamente plantas, desde el punto de vista trófico actúan como consumidores primarios. Y por supuesto ya debes estar pensando en las plantas carnívoras, pero en términos del flujo de energía son 100% productoras primarias. Su comportamiento carnívoro sí es relevante para su nutrición mineral, pero ese es otro tema. Probablemente te sorprenderá la diversidad de los descomponedores, ya que son muchos más que los hongos y bacterias del suelo que tal vez imaginaste. Organismos de prácticamente todo el abanico de diversidad animal obtienen toda o parte de su energía de la madera
Termitas, polillas, escarabajos, isópodos, larvas de moscas ¡e incluso peces!.
, la hojarascaLombrices, milpiés, caracoles, babosas, isópodos, colémbolos, numeroso insectos, ranas, sapos y peces. u otras formas de necromasa en distintas fases de descomposición. Estos ejemplos ilustran que las variables de estado del modelo de la Figura 2 están compuestas por una gran diversidad de organismos. Incluso, un organismo puede estar parcialmente en más de una de ellas (por definición, los omnívoros). Este es uno de los aspectos más distintivos de la visión ecosistémica, que incluso desafía nuestra intuición. Los individuos y las especies son una parte más patente de nuestra vida cotidiana y por lo tanto nos resulta difícil imaginar que “partes de una oveja” estén repartidas entre consumidores primarios y descomponedores. De hecho, hay evidencias de ovejas o ciervos depredando nidos de aves, con lo cual los rumiantes de un ecosistema pueden ser consumidores primarios, secundarios, terciarios y descomponedores.

Imagen 5. A estos bovinos los colocaríamos intuitivamente en la caja de consumidores primarios, pero en realidad allí están consumiendo plantas muertas y actuando como descomponedores. Unas horas antes, podrían haber estado pastando sobre plantas vivas, como verdaderos consumidores primarios.
Variables de control
Como recordarás, la Figura 2 no incluye a las “variables de control” que regulan el tamaño de las cajas o la velocidad de los flujos. Sin embargo, al analizar la Imagen 3 sobre unos lotes en el sur de Buenos Aires pudiste pensar en algunas variables que regulan la primera bifurcación del flujo de la Figura 2: la proporción de radiación incidente que es absorbida por los productores primarios. El caso más llamativo fue el pivote de riego, al cual se lo ve mucho más verde que otra vegetación no regada. ¿Cómo controla la disponibilidad de agua en el suelo a esa bifurcación? Fundamentalmente, la controla a través de un aumento del área foliar. Con mayor agua en el suelo aumenta el área foliar, lo cual “cierra” la pérdida de energía por reflexión (Figura 3). Esto puede cambiar en el tiempo, como cuando una lluvia moja un suelo, o en el espacio, como cuando viajas a lo largo de un gradiente de precipitación como el que muestra la parte derecha de la figura. La radiación incidente es aproximadamente la misma en toda la zona, pero al oeste llueven unos 1100 mm anuales y al este menos de 200.

Figura 3. El esquema muestra el flujo de energía en el nivel de productores primarios tal como en la Figura 2. La disponibilidad de agua es una variable de control que actúa sobre una llave que abre o cierra la salida por reflexión y, por lo tanto, cierra o abre la entrada por radiación absorbida. La imagen muestra un sector de la Patagonia con un fuerte gradiente de mayor precipitación al oeste a menor precipitación al este.
De la misma manera, la Figura 3 podría haber incluido más variables de control y más controles por variable. Pronto entenderás por qué decidimos simplificar la Figura 2 y dejar para más tarde las variables de control. La disponibilidad de agua no sólo controla la cantidad de radiación absorbida vs reflejada. También puede actuar sobre las pérdidas por calor entre la radiación absorbida y la PPB (Figura 4). El cierre de estomas no impide que se absorba radiación sino que limita la llegada de CO2 a los cloroplastos y, por lo tanto, limita la tasa fotosintética. Eso ha sido representado con una segunda flecha azul en la figura. Y la disponbilidad de nitrógeno inorgánico también actúa sobre el área foliar y la concentración de clorofila, y por lo tanto sobre la proporción de radiación absorbida, pero además actúa sobre la asimilación de CO2 porque es un insumo para sintetizar la la enzima RuBisCO que define la tasa fotosintética “aguas abajo” de la absorción de fotones por la clorofila.

Figura 4. Flujo de energía en el nivel de productores primarios tal como en la Figura 3. La disponibilidad de agua ahora controla no sólo la salida por reflexión sino también la salida como calor previa a la PPB (NA). Se agrega una nueva variable de control: la disponibilidad de N inorgánico que controla también ambas pérdidas.
Subsidios de energía
En un agroecosistema, el manejo procura conducir la energía solar fotosintéticamente activa con el fin de producir ciertos productos vegetales o animales. Por ejemplo, en un sistema pastoril destinado a producir carne, se busca reducir lo no utilizado por el ganado mediante cargas altas. También se puede reducir lo no asimilado mediante la utilización de forrajes de alta digestibilidad, o reducir lo respirado mediante sombra, abrigo, menor movimiento de los animales, o control de parásitos y enfermedades. Algo similar sucede en otros ecosistemas en los que la influencia humana aplica energía con el fin de preservar ciertas especies, controlar invasoras, prevenir incendios o fijar dunas.
Prácticamente todas estas intervenciones requieren aportes de energía que se suman a la provista por la radiación solar y por lo tanto se los conoce como subsidios de energía. La elevada productividad de las cosechas y de los rodeos se mantiene en parte mediante estos subsidios de energía. Por ejemplo, la energía destinada a las labranzas, la irrigación, la fertilización, la selección genética o el control de plagas puede ser estimada en calorías y así sumarse como subsidio al aporte de la radiación solar. También algunos ecosistemas reciben subsidios de energía de fuentes naturales. Por ejemplo, las marismas o los valles de inundación reciben aportes de materia orgánica y nutrientes con un equivalente energético que se suma a la energía provista por la radiación solar en el propio ecosistema.
¿Cómo pueden visualizarse los subsidios de energía en los términos del modelo de la Figura 2? En la mayoría de los casos, los subsidios son un tipo de variable de control. Por lo tanto, se los puede representar actuando sobre una o más pérdidas del flujo. Por ejemplo, el riego es un subsidio que aumenta la productividad primaria porque reduce las pérdidas por radiación reflejada y calor de los productores primarios (Figuras 3 y 4). Similar razonamiento se puede aplicar con bastante éxito a todas las intervenciones humanas. En la gran mayoría de los casos, los subsidios de energía no se incorporan al producto final (ni una kilocaloría del grano de soja proviene de una kilocaloría de gasoil o de glifosato). Las kilocalorías de gasoil o de glifosato simplemente resultaron en que se condujera mayor cantidad de energía solar a la producción de granos. Por supuesto, hay excepciones: la iluminación artificial de invernáculos y la importación de alimentos para el ganado, por ejemplo, son subsidios de energía que no actúan sobre una pérdida sino que se incorporan directamente al flujo desde una fuente externa.
Un cultivo de maíz sirve como ejemplo de flujo de energía a partir de la radiación solar (Andrade, 1998), al cual se le ha incorporado la energía de subsidio a partir de datos de un cultivo similar en Estados Unidos (Pimentel y Dazhong, 1990, Figura 5). En primer lugar, llama la atención la gran magnitud de las pérdidas por radiación reflejada, no asimilada y respiración. Las pérdidas por radiación reflejada corresponden mayoritariamente al período de barbecho, en el cual el cultivo no está presente, y a los períodos de crecimiento y senescencia, en los cuales el cultivo no llega a absorber toda la radiación incidente. Las pérdidas de energía no asimilada y respiración son proporcionalmente mayores a la recientemente descripta. Sin embargo, estas “pérdidas” representan en buena medida mecanismos importantes de productividad, como la respiración del cultivo. En segundo lugar, como consecuencia de lo anterior, es notable la pequeña proporción de energía obtenida en el grano en relación con la energía incidente (aproximadamente 0.8 % si se considera la radiación fotosintéticamente activa incidente, ó 0.4 % si se considera toda la energía incidente). Finalmente, esta cuantificación resalta la pequeñez de los subsidios de energía en comparación a los ingresos de energía solar (85 vs 25500), pero, por otro lado, también ilustra que ese comparativamente pequeño subsidio transforma lo que de otra manera hubiera sido un sistema natural en uno que produce granos. Estas dos caras de los subsidios de energía, la de su pequeñez en relación con el aporte de la naturaleza y la de su grandeza en relación con la transformación que logran, deberían ser muy aleccionadoras para quienes manejan los ecosistemas.
Aquí puedes investigar sobre subsidios de energía o “La luz que no puedes ver”.

Figura 5. Flujo de energía en un cultivo de maíz en Balcarce, Provincia de Buenos Aires. Las flechas corresponden a los flujos de la Figura 2. Las “válvulas” representan la acción de variables de control. Los números corresponden a Giga Joules/ha.año. RI indica la radiación fotosintéticamente activa incidente. Los datos del cultivo fueron tomados de Andrade (1998) y los de subsidios de energía de Pimentel y Dazhong (1990). Nótese que el flujo se ha simplificado y no incluye otros niveles tróficos.
Eficiencias
El flujo de energía, con sus diversos componentes, ingresos, pérdidas y transferencias, lleva casi naturalmente al concepto de eficiencia. En términos generales, una eficiencia es un cociente entre producto e insumo (expresada en proporción o en porcentaje). En el marco del flujo de energía de un ecosistema se pueden definir varias eficiencias relevantes para distintos propósitos. En el ejemplo de la Figura 6, el cociente entre productividad secundaria y primaria aérea equivale a 1.9% (2.6/135). De manera similar se pueden calcular otras eficiencias, como la eficiencia del uso de la radiación, una medida muy frecuentemente evaluada en sistemas agropecuarios, que consiste en el cociente entre la productividad primaria neta y la radiación absorbida (se suele tomar sólo la productividad aérea). Los subsidios de energía aumentan la eficiencia de transferencia de energía al disminuir las pérdidas y, por lo tanto, aumentar el producto para una misma cantidad de insumo. Sin embargo, puede ser de interés estimar otra eficiencia, que incluya también a la energía del subsidio como parte del insumo. Este nuevo cálculo puede resultar en una disminución de la eficiencia como consecuencia de la aplicación del subsidio. Tal vez puedas pensar en un ejemplo a partir de la Figura 5.

Figura 6. Flujo de energía parcial y simplificado de un establecimiento de La Madrid, provincia de Buenos Aires. Las flechas corresponden a los flujos de la Figura 2. Los números corresponden a Giga Joules/ha.año, excepto por los dos números recuadrados en cursiva que corresponden a variables de estado en Giga Joules/ha.año. Los datos de producción de carne y de carga fueron provistos por Fernando Pacín. Los datos de radiación en el compartimiento del forraje fueron obtenidos por el Sistema de Seguimiento Forrajero (Segf) del Laboratorio de Análisis Regional y Teledetección, FAUBA (Grigera et al. 2007a, 2007b).
Variaciones del flujo de energía entre ecosistemas
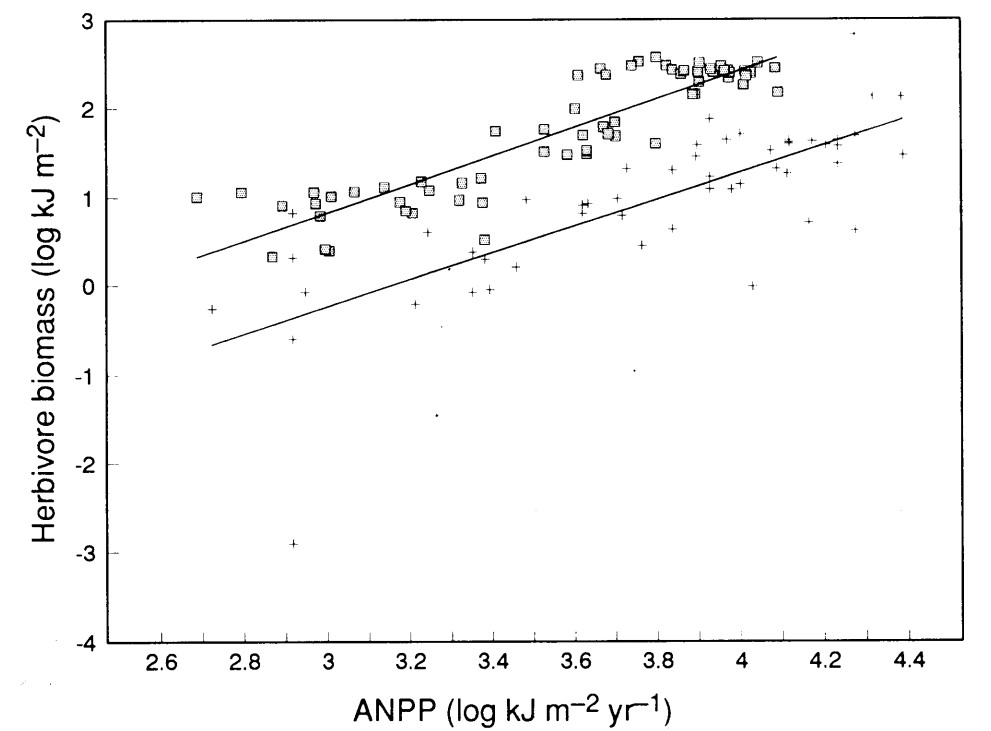
Figura 7. Biomasa de herbívoros en función de la productividad primaria neta aérea en ecosistemas naturales de diferentes lugares del mundo (cruces, línea inferior) y en sistemas ganaderos de Argentina y Uruguay (cuadrados, línea superior). El segundo grupo corresponde a departamentos provinciales en los cuales la ganadería se basa mayoritariamente en pastizales naturales. Fuente: Oesterheld et al. 1992.
A esta altura, ya deberías dominar con soltura los fundamentos del flujo de energía en los ecosistemas. Llega el momento entonces de descubrir que si bien el modelo de la Figura 2 es válido como base para cualquier ecosistema, hay grandes variaciones. El flujo de energía difiere entre ecosistemas y también a lo largo del tiempo o en en respuesta a disturbios.
Quizás el contraste más fuerte de flujo de energía entre ecosistemas es entre los terrestres y los acuáticos (una gran simplificación porque hay enorme variedad dentro de estas categorías). Los principales productores de los sistemas terrestres suelen ser las plantas superiores, mientras que en los acuáticos suelen ser los que componen el fitoplancton, un grupo diverso de organismos fotosintéticos microscópicos (p.ej., algas verdes, diatomeas, cianobacterias). Por lo general, los sistemas terrestres tienen estructuras tróficas más complejas que los acuáticos. Y el contraste más llamativo es lo que sucede con la disponibilidad de energía solar y nutrientes entre estos tipos de ecosistemas. Los sistemas acuáticos dependen de los nutrientes disueltos en el agua, los cuales suelen estar en baja disponibilidad, mientras que los terrestres los tienen en el suelo, en mayor disponibilidad. Las plantas terrestres absorben luz en el canopeo y nutrientes por las raíces en el suelo, mientras que el fitoplancton depende de la luz que se va extinguiendo con la profundidad y la concentración de nutrientes en esa profundidad.
Otro de los contrastes fuertes lo genera la variación de la magnitud relativa de los flujos de consumo y no utilizado por los herbívoros (Figura 2). Algunos ecosistemas tienen una gran carga de herbívoros en relación con su productividad primaria neta. En consecuencia, una gran proporción de la productividad primaria se destina al crecimiento y respiración de ese nivel trófico y una proporción mucho menor sigue el camino del no utilizado, también llamado vía de los detritos. En cambio, otros tienen una baja carga de herbívoros, ya sea por controles naturales o por efecto humano, y una gran proporción de la productividad primaria neta sigue el camino de la necromasa.
Un patrón bastante consistente cuando se comparan ecosistemas es que los de mayor productividad primaria neta tienen una mayor carga de herbívoros, tanto artrópodos como vertebrados (Figura 7). La figura abarca sistemas naturales desde desiertos a bosques húmedos y sistemas ganaderos, desde las estepas y semidesiertos de la Patagonia hasta los pastizales húmedos de Corrientes o de Uruguay. Las relaciones doble logarítmicas tienen pendiente mayor que 1, lo cual significa que la biomasa de herbívoros crece más pronunciadamente que la productividad primaria. Es decir, la carga por unidad de productividad primaria crece a medida que los ecosistemas son más productivos. Y la posición de la línea que representa al patrón de los ecosistemas ganaderos, tan por encima y paralela a la de los sistemas naturales, significa por un lado que en los ganaderos hay 10 veces más carga que en los naturales y, por otro, que la forma en que responden al aumento de productividad primaria es similar.
Si conectas estas observaciones con la Figura 2, verás que “simplemente” hemos puesto una lupa sobre una de las tantas bifurcaciones de energía que muestra la figura, la que define los destinos inmediatos de la productividad primaria neta. Esa bifurcación cambia a lo largo de gradientes de productividad, probablemente por múltiples razones: el clima más benigno favorece a las poblaciones de animales, la productividad primaria se hace más nutritiva y digerible con el aumento de la productividad, los humanos, desde el Pleistoceno, hemos extinguido o reducido sensiblemente a las poblaciones de herbívoros, los depredadores controlan a las poblaciones de herbívoros, etc. Puedes explorar por tu cuenta o llevar preguntas a clase sobre variaciones de otras bifurcaciones de interés dentro del modelo de la Figura 2 o de los datos comparativos de la Tabla 1.
Tabla 1. Productividad, respiración y consumo heterotrófico de distintos ecosistemas (GigaJoules/ha.año) y porcentaje de la PPN y la PNE en relación a la PPB.
| Campo de alfalfa para heno (EEUU) | Plantación joven de pino (Inglaterra) | Bosque de edad media de pino y roble (N.York) | Manantial (Silver Springs, Florida) | Selva (Puerto Rico) | Sonda Costera (L. Island, N.York) |
Productividad primaria bruta (PPB) | 1022 | 511 | 482 | 872 | 1886 | 239 |
Respiración productores (Rp) | 385 | 197 | 268 | 503 | 1341 | 134 |
Productividad primaria neta (PPN) | 637 | 314 | 210 | 369 | 545 | 105 |
Respiración heterotrófica (Rh+Rc+Rd) | 34 | 193 | 126 | 285 | 545 | 105 |
Productividad neta del | 603 | 122 | 84 | 84 | Muy baja o nula | Muy baja o nula |
Razón PPN/PPB (porcentaje) | 62,3 | 61,5 | 43,5 | 42,5 | 28,5 | 43,8 |
Razón PNE/PPB (porcentaje) | 59 | 23,8 | 17,4 | 9,6 | 0 | 0 |
Conclusiones
La energía fluye a través de los ecosistemas como resultado de interacciones complejas entre organismos y su entorno físico, desde los productores primarios hasta los descomponedores. El flujo sucede entre compartimientos que se asocian con el concepto de nivel trófico, a los cuales se agrega un reservorio de necromasa. Diversos factores abióticos y bióticos actúan como variables de control y, por lo tanto, influyen sobre la eficiencia de las transferencias de energía. En este contexto, el manejo humano y los subsidios de energía pueden modificar el flujo con diversos fines, como, por ejemplo, maximizar la productividad.
Problemas
- Productividad Primaria Bruta (PPB): 10,000 kcal/m²/año
- Respiración de los autótrofos: 4,000 kcal/m²/año
- Consumo de herbívoros: 1,500 kcal/m²/año
- Energía no utilizada por los herbívoros: 3,000 kcal/m²/año
Con base en estos datos, responde:
a. ¿Cuál es la Productividad Primaria Neta (PPN) del ecosistema?
b. Dados estos flujos, ¿qué debería suceder con la caja de los productores al final del año.
c. ¿Con qué eficiencia consumen productividad primaria neta los herbívoros?
2. La productividad neta del ecosistema (PNE) se define como la cantidad de energía que queda disponible en un ecosistema después de que se han restado todas las pérdidas por respiración de los autótrofos (plantas) y los heterótrofos (consumidores primarios, secundarios, etc.). Es decir, es la energía que puede ser acumulada en forma de biomasa o necromasa.
En un bosque templado se midieron los siguientes componentes del flujo de energía durante un año:
- Productividad Primaria Bruta (PPB): 12,000 kcal/m²/año
- Respiración de los autótrofos: 5,000 kcal/m²/año
- Respiración de los heterótrofos: 2,500 kcal/m²/año
Con base en estos datos, responde:
a. ¿Cuál es la Productividad Neta del Ecosistema (PNE)?
b. Explica la diferencia entre PPN y PNE en el contexto del ecosistema dado.
c. Si el cálculo de la PNE diera un resultado negativo durante varios años consecutivos, ¿qué implicaciones tendría sobre el ecosistema?
3. Esquematice el diagrama del flujo de energía en un cultivo de maíz en Pergamino. Identifique y describa las variables de estado, los flujos (incluyendo la cosecha) y las variables de control.
a. Indique en el diagrama los flujos que se modificarían más directamente por la aplicación de subsidios de energía para aumentar el flujo de productividad primaria no utilizada por los herbívoros, y cuáles podrían ser dichos subsidios.
b. ¿Qué modificaciones introduciría en el diagrama de flujo de energía si el modelo va a ser utilizado para evaluar el efecto de la aplicación de herbicidas sobre el rendimiento del cultivo?